Capítulo 9 Endogamia

Figura 9.1: O casamento entre nobres foi usado como um método de formar alianças políticas entre as elites. Com o tempo, devido ao número relativamente limitado de consortes em potencial, o pool gênico de muitas famílias governantes tornou-se progressivamente menor, até que todos as realezas européias estivessem relacionadas.
Ceballos & Álvarez 2013. doi: 10.1038/hdy.2013.25
Vilas et al. 2019. doi: 10.1080/03014460.2019.1687752
9.1 Endocruzamentos \(\times\) exocruzamentos
Endogamia é a produção de descendentes do cruzamento de indivíduos ou organismos relacionados geneticamente (endocruzamentos), o que leva a um aumento da frequência de genótipos homozigotos em relação a frequência esperada em cruzamentos aleatórios.
A homozigose de alelos raros (a maioria recessivos) pode levar ao aparecimento de traços deletérios (viz., depressão endogâmica).

Figura 9.2: Endocruzamentos aumentam as chances de encontro de alelos raros deléterios.
9.2 Causas de endogamia
9.2.1 Endogamia intencional
9.2.1.1 Estoques de laboratório
O endocruzamento sucessivo de animais tem sido usado para estabelecer linhagens genéticas “puras” para a pesquisa científica. Os experimentos conduzidos nesses assuntos são valiosos porque a variação genética não pode distorcer os resultados.
Rader 2004. ISBN: 9780691016368
Taft et al. 2006. doi: 10.1016/j.tig.2006.09.010
9.2.1.2 Estoques para melhoramento animal e vegetal
Em animais domésticos, o endocruzamento geralmente resulta em uma troca em que uma característica desejável é ampliada às custas de outra.
Por exemplo, a endogamia de gado leiteiro holandês (Holstein-Frísia) levou ao aumento da produção de leite, mas as vacas são mais difíceis de criar.
Zenger et al. 2007. doi: 10.1111/j.1365-2052.2006.01543.x link
Utsunomiya et al. 2013. doi: 10.1371/journal.pone.0064280 link
Mrode et al. 2019. doi: 10.1371/journal.pone.0064280 link
9.3 Freqüências genotípicas sob endogamia
Considere as sucessivas gerações de autofertilização em uma linhagem híbrida, cujas frequências alélicas iniciais \(p\) e \(q\) sejam idênticas.
As frequências genotípicas não estarão em equilíbrio de Hardy-Weinberg pois ao longo de algumas gerações irá existir uma deficiência de heterozigotos e um excesso de homozigotos.
Após \(n\) gerações de autofertilização, teremos:
- A frequência dos heterozigotos será
\[\frac {1}{2^n}\]
- A frequência dos homozigotos será \(\frac {1}{2^n}\); e
\[\frac {1- \frac {1}{2^n}}{2}\]

Figura 9.3: Endocruzamentos aumentam as chances de encontro de alelos raros deléterios.
9.4 Coeficiente de endogamia
A quantidade de endogamia em uma população pode ser medida pela redução na heterozigosidade.
O Coeficiente de Endogamia (\(F\)) compara a proporção real de genótipos heterozigotos (\(H_{Obs}\)) com proporção genótipos heterozigotos esperada caso a população cruze aleatoriamente (\(H_{Esp}\)), de forma que:
\[\begin{equation} F=\frac{H_{Esp} - H_{Obs}}{H_{Esp}} \tag{9.1} \end{equation}\]
que é o mesmo que:
\[F=1 - \frac{H_{Obs}}{H_{Esp}}\]
O coeficiente de endogamia (\(F\)) pode ser interpretado como a probabilidade de que dois alelos em um determinado locus sejam aleatoriamente idênticos por descendência (IBD).
Não há endogamia …
… quando a população se reproduz ao acaso, pois \(H_{Obs}=H_{Esp}\), e assim:
\[F=0\]
Há endogamia completa …
… quando não há heterozigotos na população, pois \(H_{Obs}=0\), e assim:
\[F=1\]
Há endogamia parcial …
… quando há heterozigotos na população, mas menos do que o esperado ao acaso, pois \(H_{Obs}\ne0\) e \(H_{Obs}<H_{Esp}\), e assim:
\[0<F<1\]
Também não há endogamia …
… quando há excesso de heterozigotos na população, pois .
9.5 Freqüências genotípicas sob endogamia
Assumindo que, para um locus dialélico, que \(H_{Esp} = 2pq\), podemos então definir a frequência real do genótipo heterozigoto em uma população endogâmica nos termos do coeficiente de endogamia, de forma que:
\[F = \frac {H_{Esp} - H_{Obs}}{H_{Esp}} = \frac {2pq - H}{2pq}\] Assim, temos que:
\[f(Aa) = H = 2pq - 2pqF\]
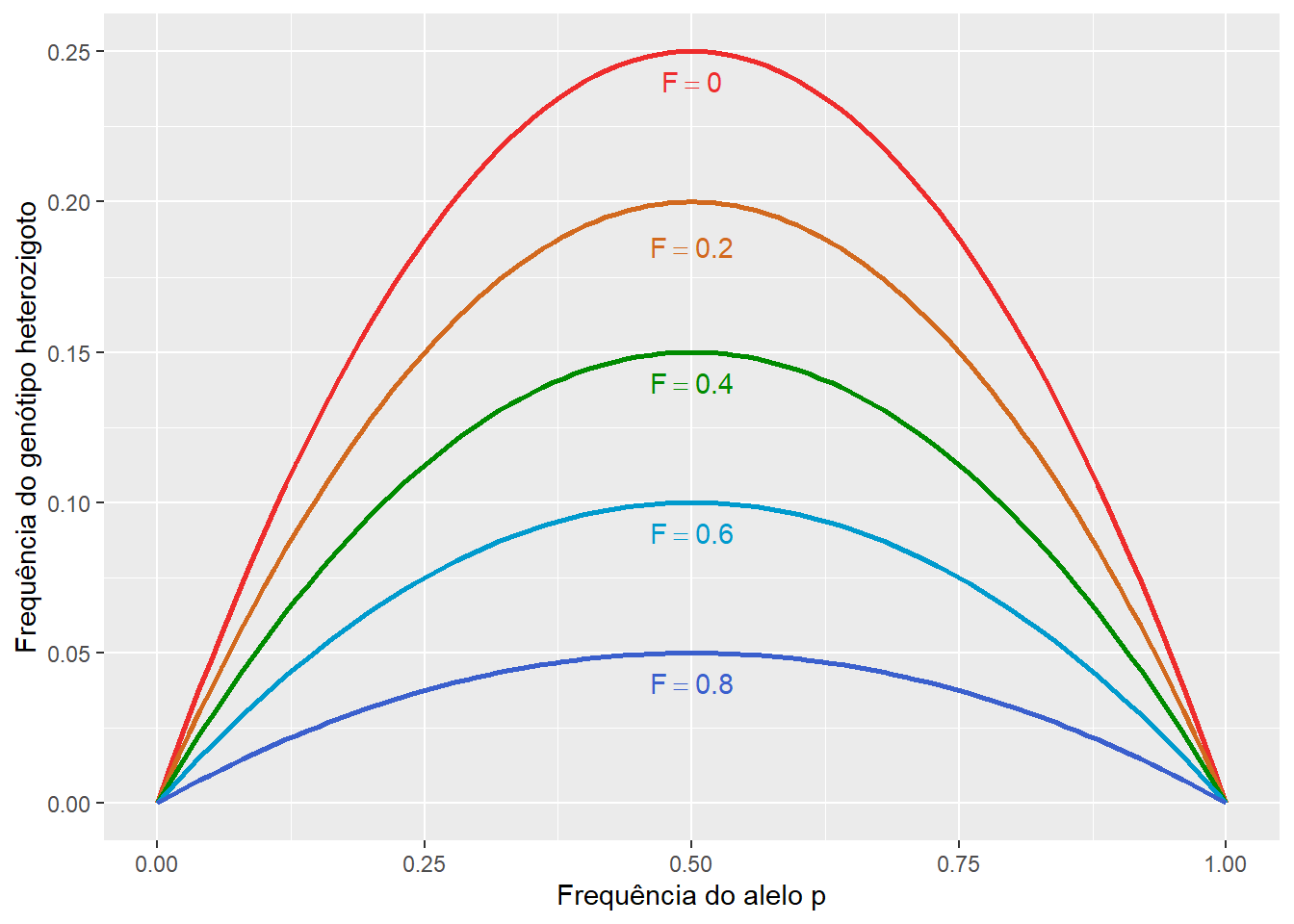
Figura 9.4: Frequência real do genótipo heterozigoto em uma população endogâmica.
Perceba que a frequência do genótipo heterozigoto sob endogamia é descrescida em uma parcela \(2pqF\) da expectativa \(2pq\) de cruzamento aleatório, parcela essa que, como visto anteriormente, será acrescida nos procionalmente aos genótipos homozigotos.
Isso pode ser demonstrado assumindo a frequência real de heterozigotos em uma população endogâmica a partir do cálculo das frequências alélicas.
Assim, teremos para o alelo \(A\):
\[ f(A) = p = D + \frac{1}{2}H = p^2 + \frac{1}{2}(2pq - 2pqF)\]
A fórmula acima pode ser reescrita, assumindo que \(pq=p(1-p)=p-p^2\), de forma que:
\[f(AA) = p^2 = p - pq + pqF = p^2 +pqF\]
E para o alelo \(a\):
\[ f(a) = q = R + \frac{1}{2}H = q^2 + \frac{1}{2}(2pq - 2pqF)\]
E assumindo que \(pq=q(1-q)=q-q^2\), de forma que:
e
\[f(aa) = q^2 = q - pq + pqF = q^2 +pqF\]
9.6 Autozigotos e alozigotos
Na fecundação, podemos categorizar os zigotos em autozigotos ou alozigotos a depender da origem dos alelos em um locus gênico.
Autozigotos — formado por dois alelos que se originam de um ancestral comum por meio de acasalamento não-aleatório, i.e., idênticos por descendência (IBD).
Alozigotos — formado por dois alelos que provêm de diferentes fontes (até onde a descendência pode ser estabelecida), ou idênticos por estado (IBS).
A chance de formação de autozigotos em uma geração depende da chance de encontros de gametas no pool gênico, que intrinsecamente depende do tamanho da amostra, de forma que:
\[\begin{equation} P(Autozigotos)=\frac{1}{2N_e} \tag{9.2} \end{equation}\]
Todos os demais cruzamentos resultam em alozigotos, sejam eles heterozigotos ou homozigotos idênticos por estado, de forma que:
\[\begin{equation} P(Alozigotos)= 1-\left(\frac{1}{2N_e}\right) \tag{9.3} \end{equation}\]

Figura 9.5: Formação de homozigotos autozigotos e alozigotos no pool gênico. Somente alelos idênticos por descendência (IBD, vermelho) formam autozigotos, uma vez que alelos idênticos por estado (IBS, amarelo) formam alozigotos, assim como os heterozigotos (verde).

Figura 9.6: Frequências genotípicas a partir das chances de formação de alozigotos e autozigotos dado o coeficiente de endogamia \(F\) da população.
9.7 Endogamia em genealogias
O coeficiente de endogamia também pode ser utilizado para expressar a probabilidade de encontro de alelos idênticos por descendência (IBD) em uma prole de endocruzamento em genealogias.
Assim, o coeficiente de endogamia de um indivíduo \(X\) da genealogia é dado pelo grau de parentesco entre seus parentais até o(s) ancestral(is) em comum, de forma que:
- Se o(s) ancestral(is) em comum não é(são) endogâmico(s):
\[F_X = \sum \left( \frac{1}{2} \right)^n\]
Onde \(n\) é o grau de separação entre os parentais até o ancestral em comum (ver Figuras 9.7 e 9.8).
- Se o(s) ancestral(is) em comum é(são) endogâmico(s):
\[F_X = \sum \left( \frac{1}{2} \right)^n \cdot \ \left( 1 + F_A \right)\]
Onde \(F_A\) é o coeficiente de endogamia do ancentral comum.

Figura 9.7: O coeficiente de endogamia (\(F\)) de um indivíduo é determinado pela chance de identidade por descendência (IBD) de genes herdados de um ou mais ancestrais comuns na sua genealogia.

Figura 9.8: Diferentes coeficientes de endogamia (\(F\)) em função das relaçoes de parentesco entre indivíduos em uma genealogia.
9.8 Endogamia ao longo das gerações
Como visto anteriormente, a chance de formação de autozigotos em uma geração depende do tamanho efetivo da população (\(N_e\)).
Seja \(\Delta F\) a variação no coeficiente de endogamia de uma geração para a outra, temos que:
\[\Delta F = \frac{1}{2N_e}\] Assim, o coeficiente de endogamia na geração atual \(t\) seria dado pela endogamia produzida na formação de autozigotos nessa e pela endogamia que já existia na geração anterior \(t-1\) herdada pelos alozigotos, de forma que:
\[F_t = \frac{1}{2N_e} + \left( 1- \frac{1}{2N_e} \right)F_{t-1}\]
ou
\[F_t = \Delta F + \left( 1- \Delta F \right)F_{t-1}\]
Assim, o coeficiente de endogamia na geração seguinte \(t+1\), pode ser expresso como:
\[F_{t+1} = \Delta F + \left( 1- \Delta F \right)F_{t}\]
As equações acima, aplicadas de forma recorrente, podem ser usadas para estimar o coeficiente de endogamia produzido em função do número de gerações \(t\) decorridas, de forma que:
\[F_t = 1 - \left( 1- \frac{1}{2N_e} \right)^t\]
Desta forma, dado tempo suficiente, i. e., \(t \rightarrow \infty\)), todas as populações finitas, irão eventualmente se tornar completamente endogâmicas.
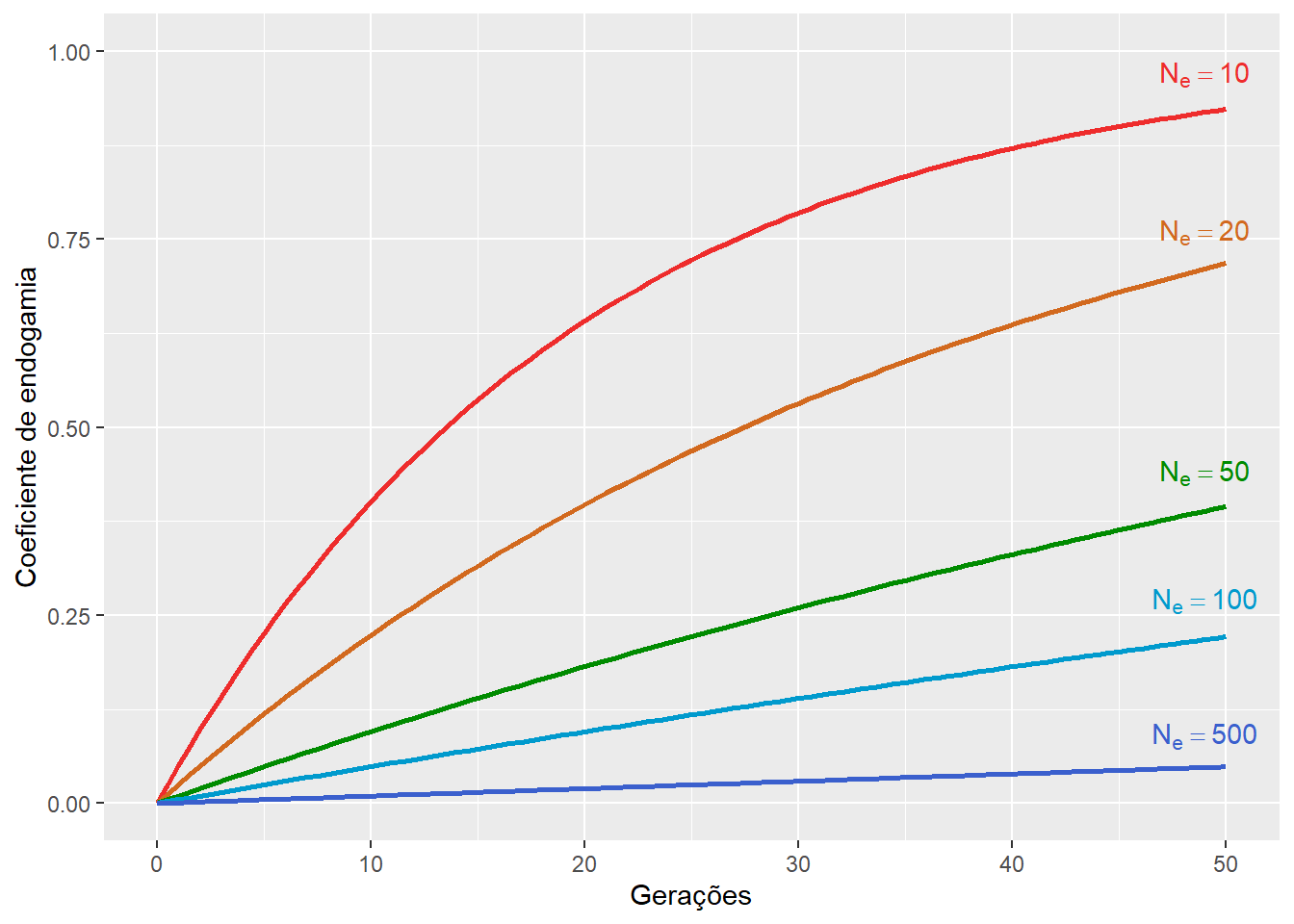
Figura 9.9: O coeficiente de endogamia ao longo das gerações em populações com diferentes tamanhos populacionaisFrequência real do genótipo heterozigoto em uma população endogâmica.
9.9 Depressão endogâmica
A depressão endogâmica é a redução da aptidão de uma determinada população devido à endogamia, a qual envolve dois mecanismos.
O aparecimento de características desvantajosas através do encontro de alelos recessivos deletérios na descendência de um cruzamento endogâmico; e
O aumento da aptidão dos heterozigotos.
Quando dois indivíduos relacionados se acasalam, a probabilidade de alelos recessivos deletérios emparelhados com a prole resultante é maior quando comparada a quando os indivíduos não relacionados se acasalam.
Assim, é possível estimar o risco relativo de um indivíduo em herdar alelos idênticos por descendência, dada simplesmente a razão entre a frequência do genótipo homozigoto dado o coeficiente de endogamia da população e a a frequência genotípica esperada sob cruzamento aleatório.
Considerando que a estimativa de risco relativo é predominantemente utilizada para genes recessivos, temos que:
\[{Risco \ Relativo} = \frac{ q^2 \left( 1-F \right) + qF }{ q^2 }\]
Por exemplo, na população humana, foi demonstrado que a taxa de mortalidade infantil é 4% maior em nascimentos de pais que são primos de primeiro grau do que em nascimentos de pais não aparentados, enquanto em aves, O fracasso da eclosão dos ovos tende a aumentar com o coeficiente de endogamia dos pais (Figura /(ref?)(fig:inbreedingdepression))
) e pássaros _Parus major_ (direita; [Noordwijk et al. 1981](https://doi.org/10.1007/BF00349183)).](figs/inbreeding_depression.svg)
Figura 9.10: Depressão endogâmica em humanos (esquerda; Bittles & Neel 1994) e pássaros Parus major (direita; Noordwijk et al. 1981).
9.10 Evitando a endogamia
A hipótese da evitação da endogamia postula que certos mecanismos se desenvolvem dentro de uma espécie, ou dentro de uma dada população de uma espécie, como resultado da seleção natural e sexual, a fim de prevenir a reprodução entre indivíduos relacionados naquela espécie ou população.
Embora a endogamia possa impor certos custos evolucionários, a abstenção de endogamia, que limita o número de parceiros potenciais para um determinado indivíduo, pode infligir custos de oportunidade.
9.10.1 Reconhecimento de parentesco
O reconhecimento de parentesco é o mecanismo pelo qual os indivíduos identificam e evitam o acasalamento com membros intimamente relacionados.
Por exemplo, os genes do complexo principal de histocompatibilidade (MHC) podem e são usados para discriminar entre indivíduos aparentados e não-relacionados
Desta forma, esses mecanismos favorecem acasalamentos heteroespecíficos.
9.10.2 Dispersão
A dispersão é adotada por algumas espécies como forma de separar os parentes próximos e evitar a endogamia.
Contudo, movimentos de longa distância podem acarretar riscos de mortalidade e custos energéticos.
9.10.3 Maturação sexual tardia
A maturação sexual tardia da prole na presença dos pais é outro mecanismo pelo qual os indivíduos evitam a endogamia.
Cenários de maturação atrasada podem envolver a remoção do genitor original do sexo oposto, como é o caso de leoas que exibem estro mais cedo após a substituição de seus pais por novos machos.
9.10.4 Cópulas extra-par
Em várias espécies, as fêmeas se beneficiam ao acasalar com vários machos, produzindo assim mais descendentes de maior diversidade genética e potencialmente de qualidade.
Fêmeas que estão ligadas a um macho de baixa qualidade genética, como pode ser o caso de endogamia, são mais propensas a se envolver em cópulas extrapar para melhorar seu sucesso reprodutivo e a capacidade de sobrevivência de sua prole.
Pusey & Wolf 1996. doi: 10.1016/0169-5347(96)10028-8
Manning et al. 1992. doi: 10.1038/360581a0
Szulkin & Sheldon 2008. doi: 10.1098/rspb.2007.0989
O’Riain et al. 2000. doi: 10.1007%2Fs002650000249
Petrie & Kempenaers 1998. doi: 10.1016/S0169-5347(97)01232-9
9.11 Exercícios
9.11.1 Exercício 1 - Coeficiente de endogamia
Considere as três populações abaixo.
| \(A_{1}A_{1}\) | \(A_{1}A_{2}\) | \(A_{2}A_{2}\) | |
|---|---|---|---|
| População 1 | 125 | 250 | 125 |
| População 2 | 50 | 30 | 20 |
| População 3 | 100 | 500 | 400 |
Qual o coeficiente de endogamia (\(F\)):
Na população 1?
Na população 2?
Na população 3?
Qual população apresenta maior nível de endocruzamento?
9.11.2 Exercício 2 - Endogamia na genealogia
O heredograma abaixo mostra um indivíduo X produzido por um cruzamento de “meio-primos de terceiro grau” (os pais são primos completos pela parte materna, e meio-primos pela parte paterna).

Figura 9.11: Heredograma hipotético da paciente X.
Calcule o coeficiente de endogamia da paciente X, considerando que:
Nenhum dos ascedentes diretos é fruto de um casamento cosanguíneo.
Seu avô A, por parte de mãe, é filho de um casamento entre primos de primeiro grau.
Qual é o risco relativo de X apresentar uma doença autossômica recessiva, como anemia falciforme, cuja prevalência pode chegar a 1:500 indivíduos em populações africanas (Xiao & Lauschke, 2021).