Capítulo 12 Migração e Fluxo Gênico

Figura 12.1: A Grande Muralha da China tem servido como uma barreira física ao fluxo gênico entre subpopulações de plantas separadas por mais de 600 anos, como por exemplo para o damasqueiro tibetano (Prunus armeniaca), em floração. Foto: runchinamarathon.com/images (2014)
Su et al. 2003. doi: 10.1038/sj.hdy.6800237
12.1 Migração \(\times\) Fluxo gênico
Como visto anteriormente, quando populações se encontram estruturadas, as subpopulações irão divergir por deriva genética.
Cabe assim à migração promover o cruzamento de indivíduos de um deme com os de outro, e logo, o fluxo de genes entre as subpopulações.
Contudo, a migração faz referência geralmente à indivíduos juvenis e adultos, e não necessariamente inclui o intercruzamento de indivíduos de populações diferentes (e. g.,, pássaros migratórios).
Enquanto o fluxo gênico requer necessariamente que haja intercruzamento, o qual é também promovido por dispersão de larvas, sementes e gametas.

Figura 12.2: A migração de indivíduos entre subpopulações promove o fluxo gênico.
Migração – O movimento de membros de uma espécie para um ambiente diferente.
Fluxo gênico – A transferência da variação genética de uma população para outra.
12.2 Modelando a migração
Conside um modelo simples onde duas populações locais trocam genes ao permutar uma proporção \(m\) do total de seus indivíduos a cada geração reciprocamente.
Assim, em um lócus com dois alelos \(A\) e \(a\), teremos…
- … na população 1:
\[f(A,Pop\ 1) = p_1\]
e
\[f(a,Pop\ 1) = q_1\]
- … e na população 2:
\[f(A,Pop\ 2) = p_2\]
e
\[f(a,Pop\ 2) = q_2\]
Assim, a frequência \(p'\) do alelo \(A\) na próxima geração dependerá da fraçao \(m\) de migrantes trocada entre as duas populações, de forma que…
- … na população 1:
\[p'_1 = (1-m)p_1 + mp_2\]
- … e na população 2:
\[p'_2 = (1-m)p_2 + mp_1\]
Onde \(m\) é a fração de migrantes entre as duas populações, e \(1-m\) é a a fração de não-migrantes.

Figura 12.3: A frequência alélica \(p\) após uma geração com uma taxa de migração recíproca \(m\) entre duas populações.
A mudança nas frequências gênicas promovida pelo fluxo gênico dependerá assim da fração \(m\) de migrantes e da diferença prévia existente entre as frequências gênicas \(p_1\) e \(p_2\) das populações, de forma que…
- … na população 1:
\[
\begin{aligned}
\Delta p_1
&= p’_1 – p_1 \\
&= (1 – m)p_1 + mp_2 \ – \ p_1 \\
&= p_1 – mp_1 + mp_2 \ – \ p_1 \\
&= mp_2 \ – \ mp_1 \\
&= m(p_2 \ – \ p_1)
\end{aligned}
\]
- … e na população 2:
\[
\begin{aligned}
\Delta p_2
&= p’_2 – p_2 \\
&= (1 – m)p_2 + mp_1 \ – \ p_2 \\
&= p_2 – mp_2 + mp_1 \ – \ p_2 \\
&= mp_1 \ – \ mp_2 \\
&= m(p_1 \ – \ p_2)
\end{aligned}
\]
Desta maneira, a migração será uma força evolutiva (promeverá mudança nas frequências gênicas), quando:
\(m > 0\) – i. e. , há fluxo gênico (viz., as populações locais não estão totalmente isoladas); e
\(p_1 \neq p_2\) – i. e., há divergência genética entre as populações locais (o que quase sempre irá ocorrer, uma vez que as subpopulações são finitas e possuem uma história evolutiva prévia, viz., deriva genética).
Assim, quando \(p_1 = 0\) e \(p_2 > 0\), O fluxo gênico introduzirá nova variação genética na população 1, uma vez que \(p’_1 = mp_2\)
Neste sentido o fluxo gênico atuará localmente como um processo criativo, semelhante à mutação, mas mais expressivo do que esta, pois geralmente \(m \gg µ\) (ver taxa de mutação em [#Mutação]).
12.3 Migração ao longo do tempo
O fluxo gênico entre populações geneticamente distintas promove mudança nas frequências gênicas (evolução).
Contudo, esta mudança ocorre em uma direção específica.
Assim, se…
- …na população 1
\[ \begin{aligned} p'_1 &= (1-m)p_1 + mp_2 \\ &= p_1 - mp_1 + mp_2 \\ &= p_1 - (mp_1 - mp_2) \\ &= p_1 - m(p_1 - p_2) \end{aligned} \]
… e na população 2:
…na população 1
\[ \begin{aligned} p'_2 &= (1-m)p_2 + mp_1 \\ &= p_2 - mp_2 + mp_1 \\ &= p_2 + (mp_1 - mp_2) \\ &= p_2 + m(p_1 - p_2) \end{aligned} \]
E se chamarmos \(d\) a diferença entre as frequências gênicas das duas populações em uma dada geração, de forma que:
\[ d = p_1 - p_2\]
Então teremos que:
\[p'_1 = p_1 - md\]
e
\[p'_2 = p_2 + md\]
Assim, a diferença entre as frequências gênicas das duas populações na geração seguinte \(d'\), será:
\[
\begin{aligned}
d'
&= p’_1 \ – \ p’_2 \\
&= (p_1 - md) - (p_2 + md) \\
&= p_1 - md - p_2 - md \\
&= p_1 - p_2 - 2md
\end{aligned}
\]
Assim, se $ d = p_1 - p_2$, logo teremos que:
\[ \begin{aligned} d' &= d - 2md \\ &= d(1-2m) \end{aligned} \]
Desta forma, o fluxo gênico entre as populações, quando \(m>0\), leva à redução da diferença entre as frequências alélicas das subpopulações, proporcional a taxa de migração \(m\), uma vez que \(d'<d\).
Logo, após \(t\) gerações de migração, temos que:
\[d_t = d(1 – 2m)^t\]
Assim:
\(d_t \to 0\), quando \(t \to \infty\);
e \(d = 0\), quando \(p_1 = p_2 = 0.5\)
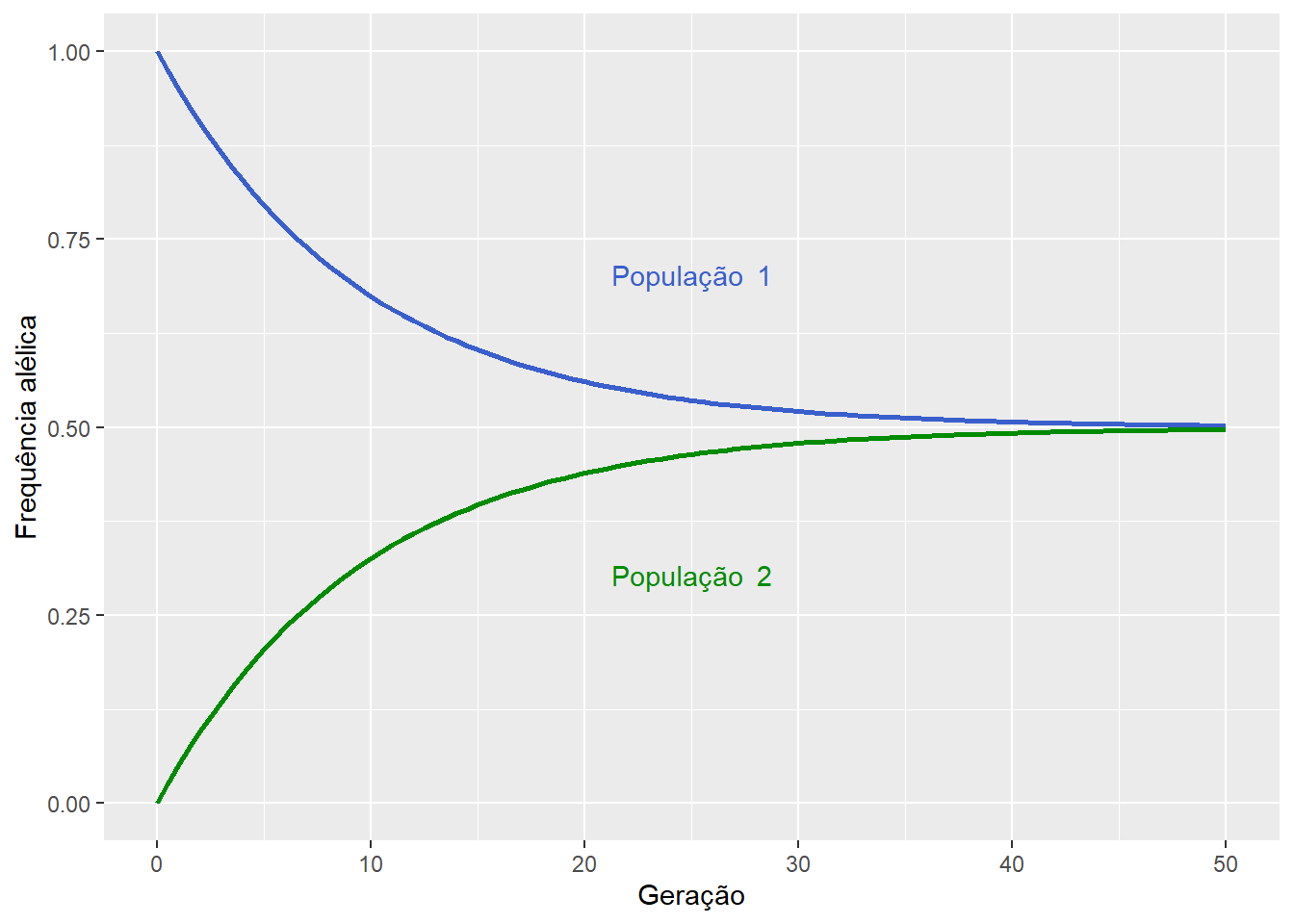
Figura 12.4: O fluxo gênico entre duas populações levará ao equilíbrio das frequências alélicas proporcionalmente a taxa de migracão \(m\).
O fluxo gênico pode ser visto como o ponto que conecta as populações:
Em curto prazo, o fluxo gênico atua como uma força evolutiva criadora, pois introduz novas variantes alélicas em populações locais divergentes ou mesmo em espécies próximas por meio de hibridização e introgressão.
Em longo prazo, o fluxo gênico atua como uma força evolutiva conservadora, pois restringe a diferenciação gênica entre populações, promovendo a conectividade e mistura.
12.4 O modelo fonte-dreno
No modelo fonte-dreno, também conhecido como continentes-ilha, o fluxo gênico ocorre unidirecionalmente, a partir de uma grande população (fonte) para uma menor (dreno).
Desta forma, a introdução de alelos vindos da população fonte afeta a freqüência alélica na população dreno, mas não ao contrário.
Como a população fonte não recebe migrantes da população dreno, ou, se o faz, a diferença no tamanho das duas populações torna a fração de migrantes desprezível, teremos que a frequência \(P\) do alelo \(A\) não é alterada pelo fluxo gênico, e permanecerá constante de uma geração para a outra.
Contudo, na população dreno, a frequência \(p\) do alelo \(A\) na próxima geração será influenciada pela fração \(m\) de migrantes oriundos da população fonte, de forma que:
\[p_1 = (1 – m)p_0 + mP\]

Figura 12.5: A frequência alélica \(p\) após uma geração com uma taxa de migração unidirecional \(m\) pa população fonte para a população dreno.
Como a frequência \(P\) não muda, podemos prever que:
\[p_2 = (1 – m)p_1 + mP\]
\[p_3 = (1 – m)p_2 + mP\]
\[...\]
E assim, em termos gerais, a frequência \(p\) do alelo \(A\) na população dreno após \(t\) gerações será:
\[p_t = (p_0 - P)(1 - m)^t+ P\]
De forma que \(p_t \to P\), quando \(t \to \infty\).
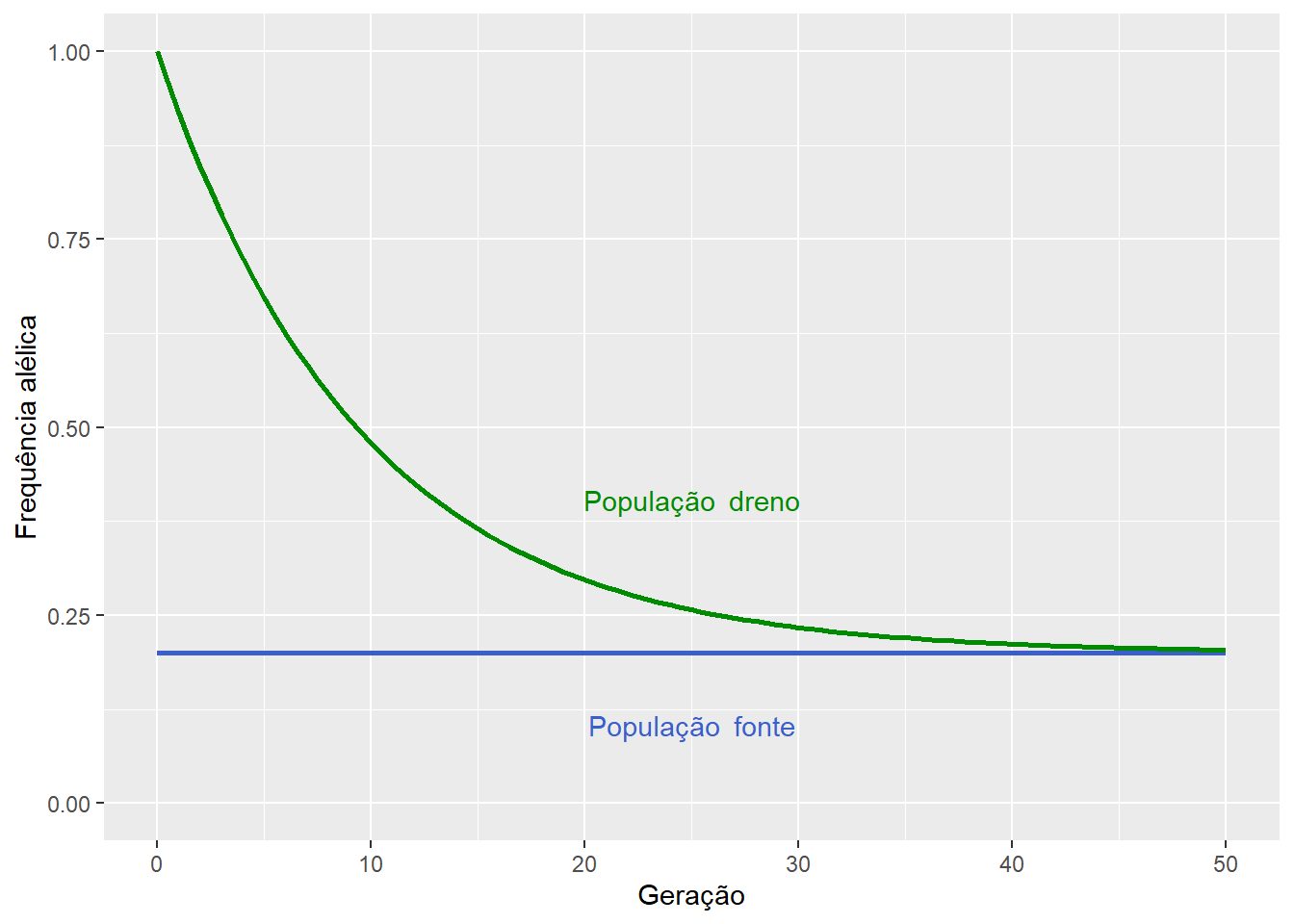
Figura 12.6: O fluxo gênico da população fonte para a população dreno promoverá, após certo tempo, dada a taxa de migracão \(m\), a substituiçào dos alelos na última.
12.5 O modelo de ilhas
O modelo de ilha é uma modelagem mais complexa da migraÇão, onde todas \(n\) populações estão conectadas através de uma taxa de migração \(m\) constante.
A maneira mais simples de modelar esse tipo de sistema é considerar que os migrantes compreendem um grande reservatório de migrantes, que são distribuídos para todas as populações.
Se \(\bar{p}\) é a frequência alélica média global, o conjunto de migrantes pode ser considerado como tendo também essa frequência alélica.
Sendo assim, a frequência \(p\) do alelo \(A\) na \(i\)-nésima população na próxima geração será:
\[p_{i,t+1} = (1-m)p_{i, t} + m\bar{p}\]
12.6 Equilíbrio entre fluxo gênico e deriva gênica
O fluxo gênico atua reduzindo a probabilidade de identidade por descendência (IBD), uma vez que um alelo introduzido de fora de uma população significa que não haverá ancestralidade comum dentro da população.
Usando o índice de fixação (\(F_{ST}\)) como a medida do progresso da deriva genética, i.e., como a probabilidade de identidade por descendência em subpopulações relativas à metapopulação total, podemos expressar assim probabilidade total de identidade por descendência em populações migrantes.
Como visto anteriormente, em uma dada geração \(t\), o coeficiente de endogamia (\(F\)) será:
\[F_t = \frac{1}{2N_e} + \left( 1- \frac{1}{2N_e} \right)F_{t-1}\]
Assumindo que a migração traz genes inteiramente não relacionados de outras populações, com probabilidade \(m\):
\((1 – m)\) – é a chance de que um alelo qualquer em uma geração não ter surgido por migração, e
\((1 – m)^2\) – é a chance de que nenhum dos dois alelos combinados em uma fecundação tenha imigrado nessa geração.
Assim, depois de contabilizar a possibilidade de migração, a probabilidade geral de identidade por descendência na geração tempo \(t\) será:
\[F_t = \left[ \frac{1}{2N_e} + \left( 1- \frac{1}{2N_e} \right)F_{t-1} \right] \left( 1-m\right)^2\]
Como o tamanho efetivo da população \(N_e\) e a taxa de migração \(m\) são os dois únicos parâmetros para determinar o coeficiente de endogamia \(F\) conforme acima, é bastante claro que os efeitos combinados da deriva genética e da migração devem tender a algum tipo de valor de equilíbrio.
Sendo o coeficiente de endogamia de equilíbrio \(\hat{F}\), onde \(\hat{F} = F_t = F_{t-1}\), teremos que:
se \(N\) e \(m\) são pequenos, a deriva supera a migração, e \(\hat{F} \approx 1\), mas
se \(N\) e \(m\) são grandes, a migração irá homogeneizar as frequências dos genes mais rapidamente do que a deriva, e \(\hat{F} \approx 0\).
E assim, o coeficiente de endogamia de equilíbrio \(\hat{F}\) será:
\[\hat{F} = \left[ \frac{1}{2N_e} + \left( 1- \frac{1}{2N_e} \right)\hat{F} \right] \left( 1-m\right)^2\]
Assintoticamente à taxa de migração \(m\), i. e., assumindo que o valor de \(m\) é pequeno o suficiente para que \(m^2 \sim 0\), teremos que:
\[ (1 - m)^2 \sim 1-2m\]
E assim, após várias simplificações algébricas da equação anterior (veja [aqui][#Provas]), teremos que:
\[\hat{F} \sim \frac{1-2m}{4N_em+1-2m} \]
Desde que \(2m \ll Nm\) e \(2m \ll 1\), teremos a relação aproximada:
\[\hat{F} \sim \frac{1}{4N_em+1} \]
Quando o índice de fixação (\(F_{ST}\)) é conhecido, podemos expressá-lo como:
\[F_{ST} \sim \frac{1}{4N_em+1} \]
e
\[N_em \sim \frac{ \frac{1}{F_{ST}}-1}{4} \]
Neste contexto, \(N_em\) representa o número efetivo de migrantes em uma população.
Por esse motivo, algumas pessoas passaram a ver estimativas de \(N_em\) como medidas de “fluxo gênico”, quando, na verdade, \(N_em\) não mede fluxo gênico, o que é dado efetivamente pela taxa de migração \(m\).
O que \(N_em\) realmente irá medir é a “tendência de produzir \(F_{ST}\) sob deriva aleatória”, desde que as suposições bastante irrealistas do modelo de ilha sejam cumpridas.
Assim:
Se \(N_em > 1\), há fluxo gênico suficiente para negar os efeitos da deriva genética, e manter a conectividade entre as populações, mas
Se \(N_em > 4\), então as populações locais podem ser consideradas pertencer a uma única população panmítica.
).](bookdownproj_files/figure-html/fixationmigration-1.png)
Figura 12.7: O índice de fixação (\(F_{ST}\)) é uma função não-linear do número efetivo de migrantes (\(N_em\)). Ver Whitlock & McCauley (1999).
12.7 Exercícios
12.7.1 Exercício 1 – Duas subpopulações quaisquer
Suponha que existam duas subpopulações, \(X\) e \(Y\), com frequencias gênicas iniciais de um alelo \(A\) iguais a \(p_{0,X}=0.2\) e \(p_{0,Y}=0.8\), respectivamente, sendo que \(1\%\) dos indivíduos em uma subpopulação em qualquer geração são migrantes.
Desta forma, calcule a frequência alélica nas duas populações após…
… 5 gerações.
… 25 gerações.
… 100 gerações.
12.7.2 Exercício 2 – Aranha do deserto
Os indivíduos de uma espécie de aranha do deserto (Agelenopsis aperta) apresentam características comportamentais de base genética que os adaptam às pressões seletivas de diferentes ambientes:
As aranhas de terras áridas, com baixa pressão de predação e baixa disponibilidade de presas, são muito territoriais e não discriminam a presa.
As aranhas de floresta ribeirinha, com alta pressão de predação por pássaros e abundância de presas, são pouco territoriais e discriminam entre itens de presa com maior ou menor retorno energético.
Indivíduos de uma população de floresta ribeirinha do Arizona que é cercada por populações de áreas áridas, apresentam uma mistura de comportamentos de áreas áridas e ribeirinhas.
Esses comportamentos de áreas áridas podem ser considerados mal-adaptados a um ambiente ribeirinho, onde os locais de teia são abundantes, as presas são abundantes e as pressões de predação são altas.
Estaria o fluxo gênico mantendo esses comportamentos mal-adaptativos na população ribeirinha do Arizona?
Usando os dados abaixo de isoenzimas isoenzimas para cinco populações locais da aranha do deserto (uma população da floresta ribeirinha e quatro populações próximas de habitats mais áridos), obtidos por Riechert (1993):
| Locus | \(F_{ST}\) |
|---|---|
| PGI1 | 0.020 |
| PGM1 | 0.023 |
| EST1 | 0.021 |
| MDH2 | 0.032 |
| IDH1 | 0.019 |
| GOT2 | 0.031 |
| αGPD1 | 0.029 |
| SOD1 | 0.145 |
| LDH1 | 0.012 |
| MDH1 | 0.141 |
| GAM1 | 0.063 |
| HEX1 | 0.034 |
| GOT1 | 0.033 |
| FUM2 | 0.129 |
O que pode-se concluir?